Ultraviolet radiation and skin cancer
|
|
|
Skin cancer incidence |
||
|
Skin cancer is the most common neoplasm in Caucasians in the United States, with a lifetime risk nearly equal to that of all other cancers combined [1]. The annual incidence of non-melanoma skin cancer (basal cell carcinoma or squamous cell carcinoma) was estimated at 900 000 to 1.2 million for the United Sates in 1994, which is up from 600 000 just 5 years earlier [2]. Although the incidence of melanoma is considerably lower, its more aggressive biological behaviour (responsible for 7500 deaths in the United States in 1994) has also been considered reason for concern [3]. |
||
|
|
|
|
Relationship between UV exposure and skin cancer |
|
Extensive clinical and epidemiological evidence indicates that exposure to the ultraviolet (UV) component of sunlight is the main causative factor in the development of skin cancers [4-7]. It is believed that decreases in stratospheric ozone (which shields the earth from short wavelength UV) [8], socio-economic factors that permit more time for outdoor leisure activities, and current societal standards which consider tanned skin attractive account (at least in part) for the recent increases in cutaneous malignancies by increasing cutaneous UV exposure [9]. |
|
|
|
The UV spectrum, types of damage, and relative cancer risk |
|
The biological effects and relative occurrence of different UV wavelengths, varies considerably across the UV spectrum. This has led to the division of the spectrum into three ranges on the basis of wavelength (summarised in table below): UV-A (320-400 nm), UV-B (280-320 nm) and UV-C (200-280 nm). UV-A (also referred to as long wave or near-UV) can penetrate as deep as the basal layer and capillary bed of exposed skin. It induces primarily oxidative DNA damage to various cellular components, however as damaged proteins and lipids are readily degraded and resynthesised, damage to DNA is thought to have the most significant consequences [10]. Although UV-A accounts for almost 95% of UV radiation to reach the surface of the earth, it accounts for less than 25% of the associated skin cancer risk [11]. However, it is believed to play a principal role in photoaging of exposed skin [12, 13]. |
|
At the other end of the spectrum is UV-C (also referred to as short wave, or far UV) which is directly absorbed by DNA and results in inappropriate bonding between adjacent pyrimidines. Although exposure to UV-C is highly carcinogenic in lab settings, it does not contribute to skin cancer in the general population as solar UV-C is almost completely absorbed by ozone in the earth's stratosphere and does not reach the surface of the earth [14]. Rather, it is UV-B (280-320 nm) that is calculated to account for greater than 75% of the skin cancer risk, even though it comprises only about 5% of terrestrial UV [11]. Although UV-B also produces oxidative intermediates, it is its ability to induce direct UV-C-like DNA damage that is responsible for its carcinogenic potential [15, 16]. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
direct photodamage |
|
|
|
| Oxidative DNA damage |
|
Probably the most abundant source of DNA alterations arises from the constant exposure to reactive oxygen species. These species are formed continually as by-products of cellular metabolism, although they can also be induced by exogenous sources such as ionising radiation, long wavelength ultraviolet radiation and H2O2. It has been estimated that every day as many as 10 000 oxidation reactions cause DNA damage in one human cell [17]. This damage includes DNA single strand and double-strand breaks, abasic sites, DNA-protein crosslinks and over 35 different base modifications [18-23]. Oxidative DNA damage has been implicated in the etiology of ageing, as well as that of cancer [24-26]. |
|
|
|
|
|
DNA photoproducts induced by UV-B and UV-C |
|
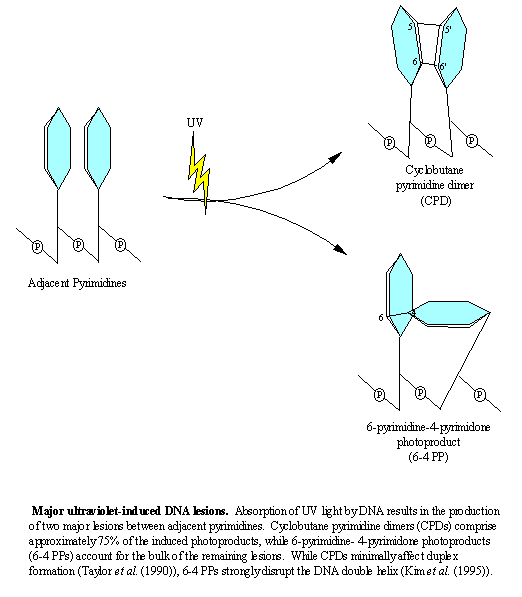
The DNA damage induced by UV-B and UV-C form two major classes of pyrimidine photoproducts: cyclobutane pyrimidine dimers (CPDs) and (6-4) pyrimidine-pyrimidone photoproducts (6-4 PP) [27]. These photoproducts are formed in DNA at approximately 3:1 proportions [28], both distort normal base-pair stacking (although this is considerably more pronounced for the 6-4 PP, resulting in a bending of the DNA by 44o compared to only 7-9o for a CPD [29-31] (see Figure)), and both have been implicated in neoplastic transformation [28, 32, 33]. |
|
|
|
The accumulation of damaged DNA has considerable consequences for living organisms. Both CPDs and 6-4 PPs (as well as some forms of oxidative base damage) can directly inhibit transcription and DNA replication processes [34-37]. When replication of damaged DNA does occur, there is an increased likelihood of misincorporating nucleotides into the nascent strand, giving rise to base substitutions [38]. |
|
|
|
However, additional alternative repair mechanisms exist in some species. |
|
|
|
|
| References |
|
1. Scotto, J., T.R. Fears, and J.F. Fraumeni, Incidence of non-melanoma skin cancer in the United States, . 1982, U.S. Department of Health and Human Services: Bethesda, MD. 2. Miller, D.L. and M.A. Weinstock, Nonmelanoma skin cancer in the United States: incidence. J Am Acad Dermatol, 1994. 30(5 Pt 1): p. 774-8. 3. Glass, A.G. and R.N. Hoover, The emerging epidemic of melanoma and squamous cell skin cancer. Jama, 1989. 262(15): p. 2097-100. 4. Vitasa, B.C., et al., Association of nonmelanoma skin cancer and actinic keratosis with cumulative solar ultraviolet exposure in Maryland watermen. Cancer, 1990. 65(12): p. 2811-7. 5. Elwood, J.M., et al., Etiological differences between subtypes of cutaneous malignant melanoma: Western Canada Melanoma Study. J Natl Cancer Inst, 1987. 78(1): p. 37-44. 6. Urbach, F., Welcome and introduction: evidence and epidemiology of ultraviolet- induced cancers in man. Natl Cancer Inst Monogr, 1978(50): p. 5-10. 7. Kligman, L.H., F.J. Akin, and A.M. Kligman, Sunscreens prevent ultraviolet photocarcinogenesis. J Am Acad Dermatol, 1980. 3(1): p. 30-5. 8. Henriksen, T., et al., Ultraviolet-radiation and skin cancer. Effect of an ozone layer depletion. Photochem Photobiol, 1990. 51(5): p. 579-82. 9. Mukhtar, H. and C.A. Elmets, Photocarcinogenesis: mechanisms, models and human health implications. Photochem Photobiol, 1996. 63(4): p. 356-7. 10. Bohr, V.A. and G.L. Dianov, Oxidative DNA damage processing in nuclear and mitochondrial DNA. Biochimie, 1999. 81(1-2): p. 155-60. 11. IARC monographs on the evaluation of carcinogenic risks to humans. Solar and ultraviolet radiation. IARC Monogr Eval Carcinog Risks Hum, 1992. 55: p. 1-316. 12. Hanson, K.M. and J.D. Simon, Epidermal trans-urocanic acid and the UV-A-induced photoaging of the skin. Proc Natl Acad Sci U S A, 1998. 95(18): p. 10576-8. 13. Ryu, A., et al., Cross-linking of collagen by singlet oxygen generated with UV-A. Chem Pharm Bull (Tokyo), 1997. 45(8): p. 1243-7. 14. Barton, I.J. and D.F. Robertson, Measurements of erythemally effective ultraviolet radiation and total ozone content. Nature, 1975. 258(5530): p. 68-9. 15. Yarosh, D., et al., Pyrimidine dimer removal enhanced by DNA repair liposomes reduces the incidence of UV skin cancer in mice. Cancer Res, 1992. 52(15): p. 4227-31. 16. Bito, T., et al., Reduction of ultraviolet-induced skin cancer in mice by topical application of DNA excision repair enzymes. Photodermatol Photoimmunol Photomed, 1995. 11(1): p. 9-13. 17. Helbock, H.J., et al., DNA oxidation matters: the HPLC-electrochemical detection assay of 8- oxo-deoxyguanosine and 8-oxo-guanine. Proc Natl Acad Sci U S A, 1998. 95(1): p. 288-93. 18. Cadet, J., et al., Oxidative damage to DNA: formation, measurement, and biological significance. Rev Physiol Biochem Pharmacol, 1997. 131: p. 1-87. 19. Henle, E.S. and S. Linn, Formation, prevention, and repair of DNA damage by iron/hydrogen peroxide. J Biol Chem, 1997. 272(31): p. 19095-8. 20. Dizdaroglu, M., Chemical determination of free radical-induced damage to DNA. Free Radic Biol Med, 1991. 10(3-4): p. 225-42. 21. Epe, B., DNA damage profiles induced by oxidizing agents. Rev Physiol Biochem Pharmacol, 1996. 127: p. 223-49. 22. McBride, T.J., B.D. Preston, and L.A. Loeb, Mutagenic spectrum resulting from DNA damage by oxygen radicals. Biochemistry, 1991. 30(1): p. 207-13. 23. Friedberg, E.C., G.C. Walker, and W. Siede, DNA repair and mutagenesis. 1995, Washington D.C.: ASM Press. 698. 24. Feig, D.I., T.M. Reid, and L.A. Loeb, Reactive oxygen species in tumorigenesis. Cancer Res, 1994. 54(7 Suppl): p. 1890s-1894s. 25. Wiseman, H., H. Kaur, and B. Halliwell, DNA damage and cancer: measurement and mechanism. Cancer Letters, 1995. 93(1): p. 113-20. 26. Beckman, K.B. and B.N. Ames, Oxidative decay of DNA. J Biol Chem, 1997. 272(32): p. 19633-6. 27. Lippke, J.A., et al., Distribution of UV light-induced damage in a defined sequence of human DNA: detection of alkaline-sensitive lesions at pyrimidine nucleoside- cytidine sequences. Proc Natl Acad Sci U S A, 1981. 78(6): p. 3388-92. 28. Mitchell, D.L. and R.S. Nairn, The biology of the (6-4) photoproduct. Photochem Photobiol, 1989. 49(6): p. 805-19. 29. Ciarrocchi, G. and A.M. Pedrini, Determination of pyrimidine dimer unwinding angle by measurement of DNA electrophoretic mobility. J Mol Biol, 1982. 155(2): p. 177-83. 30. Wang, C.I. and J.S. Taylor, Site-specific effect of thymine dimer formation on dAn.dTn tract bending and its biological implications. Proc Natl Acad Sci U S A, 1991. 88(20): p. 9072-6. 31. Kim, J.K., D. Patel, and B.S. Choi, Contrasting structural impacts induced by cis-syn cyclobutane dimer and (6-4) adduct in DNA duplex decamers: implication in mutagenesis and repair activity. Photochemistry & Photobiology, 1995. 62(1): p. 44-50. 32. Brash, D.E., et al., A role for sunlight in skin cancer: UV-induced p53 mutations in squamous cell carcinoma. Proceedings of the National Academy of Sciences of the United States of America, 1991. 88(22): p. 10124-8. 33. Protic-Sabljic, M., et al., UV light-induced cyclobutane pyrimidine dimers are mutagenic in mammalian cells. Mol Cell Biol, 1986. 6(10): p. 3349-56. 34. Protic-Sabljic, M. and K.H. Kraemer, Reduced repair of non-dimer photoproducts in a gene transfected into xeroderma pigmentosum cells. Photochem Photobiol, 1986. 43(5): p. 509-13. 35. Moore, P. and B.S. Strauss, Sites of inhibition of in vitro DNA synthesis in carcinogen- and UV- treated phi X174 DNA. Nature, 1979. 278(5705): p. 664-6. 36. Sauerbier, W. and K. Hercules, Gene and transcription unit mapping by radiation effects. Annu Rev Genet, 1978. 12: p. 329-63. 37. Htun, H. and B.H. Johnston, Mapping adducts of DNA structural probes using transcription and primer extension approaches. Methods in Enzymology, 1992. 212: p. 272-94. 38. Feig, D.I. and L.A. Loeb, Oxygen radical induced mutagenesis is
DNA polymerase specific. J Mol Biol, 1994. 235(1): p. 33-41.
|
|
|