Base excision repair (BER)
| - Short
patch BER
- Long patch BER - Consequences of defective BER - Functional overlap of NER and BER pathways |
|
|
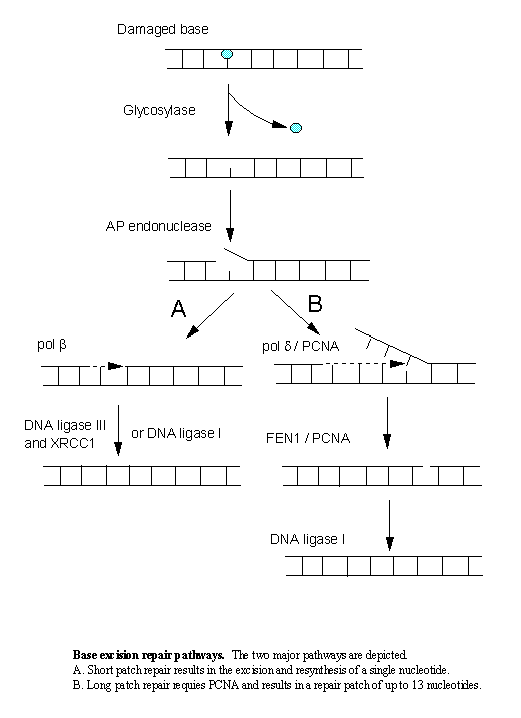
| The principal repair pathway for the removal of oxidative damage is BER. Enzymes in this pathway recognise relatively few but frequent DNA lesions such as abasic sites, deaminated C and A, and individual bases damaged by oxidative intermediates or alkylating agents. Most of these lesions do not greatly distort the DNA helix structure. In both prokaryotes and eukaryotes, BER is initiated by DNA glycosylases, a class of enzymes that each recognise a specific set of modified bases (reviewed in [1]). DNA glycosylases cleave the N-glycosylic bond between the target base and the deoxyribose, releasing a free base and leaving an apurinic/apyrimidinic (AP) site. In mammalian cells two pathways for the processing of AP sites have been described: a single nucleotide insertion (or "short patch") pathway, catalysed by DNA polymerase b [2, 3], which is almost identical to the bacterial model; and a proliferating cell nuclear antigen (PCNA) dependent ("long patch") pathway, involving a resynthesis patch of 2 to <13 nucleotides [4, 5] (see Figure). Additional variations on each of these BER pathways have also been reported (reviewed in [6, 7]). |
|
|
|
|
| Short patch BER |
|
In the "short patch" model an AP endonuclease cleaves the phosphodiester bond immediately 5' to the AP site, generating 5'-sugar-phosphate and 3'-OH ends as it nicks the DNA [8]. Removal of the 5'-sugar-phosphate moeity by a deoxyribophosphodiesterase (dRpase) results in a single nucleotide gap that is then filled by a DNA polymerase and sealed by DNA ligase [3, 9-11]. In mammalian cells DNA polymerase b (DNA pol b) is the major polymerase for this pathway [3, 9-11]. Interestingly, DNA pol b also contains intrinsic dRPase activity, and can remove the 5'-sugar-phosphate residue itself before filling in the single nucleotide gap [12, 13]. Ligation of the resynthesised nucleotide is mediated by either DNA ligase I or an XRCC1/DNA ligase III complex, which have each been shown to interact with DNA pol b [3, 14, 15]. |
|
|
| Long patch BER |
|
Although the "short-patch" pathway appears to be the most active for the repair of AP sites, an alternative "long patch" BER pathway has been reported [4, 16]. As in "short patch" BER repair, the AP sites are processed by an AP endonuclease which cleaves immediately 5' to the AP site, generating 5'-sugar-phosphate and 3'-OH ends. However, in this process, the 5'-sugar phosphate residue is not removed by a dRpase, rather DNA pol d or e adds several nucleotides to the 3' end of the nick displacing the 5'-sugar-phosphate as part of a single stranded flap structure. This flap structure is recognised and excised by flap endonuclease (FEN1) and DNA is finally ligated by DNA ligase I. This pathway is dependent on PCNA, which presumably plays a role in loading DNA pol d or e onto the DNA as well as stimulating the activity of FEN1 [17]. |
|
|
| Consequences of defective BER |
|
Oxidative DNA damage has been implicated in the etiology of ageing, as well as that of cancer [18-20]. Defects in key activities of the BER process, such as AP endonuclease, DNA pol b and the XRCC1-DNA ligase III heterodimer, lead to embryonic lethal phenotypes in mice, indicating that repair of endogenous DNA lesions is essential during embryonic development [11, 21, 22]. However, Cockayne syndrome may serve as a human model for defective BER. CS cells exhibit an impaired ability to remove oxidative damage from active genes, but not from inactive DNA [23, 24]. This suggests the existence of a mechanism coupling BER to transcription, and allowing for the preferential removal of damage in active genes, as has been demonstrated for NER [25, 26]. Indeed, CS cells appear to be defective in the transcription-coupled repair (TCR) components of both NER and BER. Furthermore there it has been demonstrated that the breast cancer gene BRCA1 plays a role in the transcription-coupled repair of oxidative damage [27, 28]. This suggests a possible link between BER capacity and the predisposition towards breast cancer. |
|
|
|
|
| Functional overlap of NER and BER pathways |
|
Further indications of an overlap between these two excision pathways, arise from the recent demonstration that the NER protein, XPG, plays a direct role role in activating a glycosylase involved in BER [29, 30], and the defects in XPG which are associated with the presentation of CS symptoms are also associated with deficiencies in BER (in addition to the NER defect associated with all XP-G individuals) [24]. Additionally, NER processes have been implicated in the repair of AP sites and a variety of single base modifications [31-36]. This has led to suggestions that, at least for some types of damage, NER may act as a backup for the BER pathway. |
|
|
|
|
| References |
|
1. Krokan, H.E., R. Standal, and G. Slupphaug, DNA glycosylases in the base excision repair of DNA. Biochem J, 1997. 325(Pt 1): p. 1-16. 2. Dianov, G., A. Price, and T. Lindahl, Generation of single-nucleotide repair patches following excision of uracil residues from DNA. Mol Cell Biol, 1992. 12(4): p. 1605-12. 3. Kubota, Y., et al., Reconstitution of DNA base excision-repair with purified human proteins: interaction between DNA polymerase beta and the XRCC1 protein. EMBO Journal, 1996. 15(23): p. 6662-70. 4. Frosina, G., et al., Two pathways for base excision repair in mammalian cells. Journal of Biological Chemistry, 1996. 271(16): p. 9573-8. 5. Klungland, A. and T. Lindahl, Second pathway for completion of human DNA base excision-repair: reconstitution with purified proteins and requirement for DNase IV (FEN1). Embo J, 1997. 16(11): p. 3341-8. 6. Wilson, S.H., Mammalian base excision repair and DNA polymerase beta. Mutat Res, 1998. 407(3): p. 203-15. 7. Bohr, V.A. and G.L. Dianov, Oxidative DNA damage processing in nuclear and mitochondrial DNA. Biochimie, 1999. 81(1-2): p. 155-60. 8. Demple, B. and L. Harrison, Repair of oxidative damage to DNA: enzymology and biology. Annu Rev Biochem, 1994. 63: p. 915-48. 9. Dianov, G. and T. Lindahl, Reconstitution of the DNA base excision-repair pathway. Curr Biol, 1994. 4(12): p. 1069-76. 10. Singhal, R.K., R. Prasad, and S.H. Wilson, DNA polymerase beta conducts the gap-filling step in uracil-initiated base excision repair in a bovine testis nuclear extract. Journal of Biological Chemistry, 1995. 270(2): p. 949-57. 11. Sobol, R.W., et al., Requirement of mammalian DNA polymerase-beta in base-excision repair. Nature, 1996. 379(6561): p. 183-6. 12. Matsumoto, Y. and K. Kim, Excision of deoxyribose phosphate residues by DNA polymerase beta during DNA repair. Science, 1995. 269(5224): p. 699-702. 13. Piersen, C.E., et al., Evidence for an imino intermediate in the DNA polymerase beta deoxyribose phosphate excision reaction. Journal of Biological Chemistry, 1996. 271(30): p. 17811-5. 14. Prasad, R., et al., Specific interaction of DNA polymerase beta and DNA ligase I in a multiprotein base excision repair complex from bovine testis. Journal of Biological Chemistry, 1996. 271(27): p. 16000-7. 15. Cappelli, E., et al., Involvement of XRCC1 and DNA ligase III gene products in DNA base excision repair. J Biol Chem, 1997. 272(38): p. 23970-5. 16. Fortini, P., et al., Different DNA polymerases are involved in the short- and long-patch base excision repair in mammalian cells. Biochemistry, 1998. 37(11): p. 3575-80. 17. Li, X., et al., Lagging strand DNA synthesis at the eukaryotic replication fork involves binding and stimulation of FEN-1 by proliferating cell nuclear antigen. J Biol Chem, 1995. 270(38): p. 22109-12. 18. Feig, D.I., T.M. Reid, and L.A. Loeb, Reactive oxygen species in tumorigenesis. Cancer Res, 1994. 54(7 Suppl): p. 1890s-1894s. 19. Wiseman, H., H. Kaur, and B. Halliwell, DNA damage and cancer: measurement and mechanism. Cancer Letters, 1995. 93(1): p. 113-20. 20. Beckman, K.B. and B.N. Ames, Oxidative decay of DNA. J Biol Chem, 1997. 272(32): p. 19633-6. 21. Xanthoudakis, S., et al., The redox/DNA repair protein, Ref-1, is essential for early embryonic development in mice. Proceedings of the National Academy of Sciences of the United States of America, 1996. 93(17): p. 8919-23. 22. Tebbs, R.S., et al., Requirement for the XRCC1 DNA base excision repair gene during early mouse development. Dev Biol, 1999. 208(2): p. 513-29. 23. Leadon, S.A. and P.K. Cooper, Preferential repair of ionizing radiation-induced damage in the transcribed strand of an active human gene is defective in Cockayne syndrome. Proceedings of the National Academy of Sciences of the United States of America, 1993. 90(22): p. 10499-503. 24. Cooper, P.K., et al., Defective transcription-coupled repair of oxidative base damage in Cockayne syndrome patients from XP group G. Science, 1997. 275(5302): p. 990-3. 25. Venema, J., et al., The genetic defect in Cockayne syndrome is associated with a defect in repair of UV-induced DNA damage in transcriptionally active DNA. Proceedings of the National Academy of Sciences of the United States of America, 1990. 87(12): p. 4707-11. 26. van Hoffen, A., et al., Deficient repair of the transcribed strand of active genes in Cockayne's syndrome cells. Nucleic Acids Research, 1993. 21(25): p. 5890-5. 27. Abbott, D.W., et al., BRCA1 expression restores radiation resistance in BRCA1-defective cancer cells through enhancement of transcription-coupled DNA repair. J Biol Chem, 1999. 274(26): p. 18808-12. 28. Gowen, L.C., et al., BRCA1 required for transcription-coupled repair of oxidative DNA damage. Science, 1998. 281(5379): p. 1009-12. 29. Bessho, T., Nucleotide excision repair 3' endonuclease XPG stimulates the activity of base excision repairenzyme thymine glycol DNA glycosylase. Nucleic Acids Res, 1999. 27(4): p. 979-83. 30. Klungland, A., et al., Base excision repair of oxidative DNA damage activated by XPG protein. Mol Cell, 1999. 3(1): p. 33-42. 31. Van Houten, B., Nucleotide excision repair in Escherichia coli. Microbiol Rev, 1990. 54(1): p. 18-51. 32. Satoh, M.S., et al., DNA excision-repair defect of xeroderma pigmentosum prevents removal of a class of oxygen free radical-induced base lesions. Proceedings of the National Academy of Sciences of the United States of America, 1993. 90(13): p. 6335-9. 33. Sancar, A., Mechanisms of DNA excision repair. Science, 1994. 266(5193): p. 1954-6. 34. Reardon, J.T., et al., In vitro repair of oxidative DNA damage by
human nucleotide excision repair system: possible
35. Otto, A.I., et al., Differential behaviors toward ultraviolet A and B radiation of fibroblasts and keratinocytes from normal and DNA-repair-deficient patients. Cancer Res, 1999. 59(6): p. 1212-8. 36. Runger, T.M., B. Epe, and K. Moller, Repair of ultraviolet B and
singlet oxygen-induced DNA damage in xeroderma pigmentosum cells. J Invest
Dermatol, 1995. 104(1): p. 68-73.
|
|
|