Mismatch repair
|
|
| Every time a cell divides it has to copy all of its DNA, so that each daughter cell can keep a complete copy. DNA replication processes are also an essential component of recombination and most DNA repair pathways. While DNA replication is highly efficient and accurate, some mistakes occasionally slip though the proof-reading apparatus, which appear as "mismatched" base pairs. Additionally, slippage of the nascent strand relative to its template during DNA replication of repeated short base sequences (known as microsatellite DNA) can lead to small loops of unpaired nucleotides. It is the process of mismatch repair (MMR) which is primarily responsible for removing these types of DNA modification (reviewed in [1-4]). In contrast to the other repair mechanisms outlined here, the distortions recognised by MMR do not contain damaged bases. It is therefor important that the repair process include a mechanism for disciminating between the two strands so that it is the newly synthesised strand which is repaired to match the parental strand, rather than mutating the parent to match the daughter. |
|
|
| Prokaryotic MMR
In E. coli a homodimer of the MutS protein, which binds to a variety of mispaired bases and small (1-5 bases) single-stranded loops, is responsible for recognition of mismatched bases [5-7] The MutS-heteroduplex DNA complex then recruits a homodimer of MutL in an ATP-dependent reaction [8]. The precise role of MutL has not yet been determined, although it has been proposed to function as a molecular matchmaker between MutS and MutH, the final component of the MMR recognition/incision complex [9]. DNA is then threaded through the complex in a bidirectional ATP-dependent manner to generate a looped structure, which has been termed the a-loop because of its similarity to the shape of that Greek letter [10]. Discrimination between the parental (or template) strand and the "flawed" daughter strand relies on the (transient) absence of adenine methylation at GATC sequences in the newly synthesised daughter strand. Since methylation lags behind replication in E. coli by several minutes, MMR is believed to occur on newly synthesised strands immediately behind the replication fork. Mut H, in the MMR complex at the base of the loop, recognises hemimethylated GATC sequences and, catalysed by MutS and MutL, introduces a single strand nick 5' to the unmethylated GATC site. The nick, which can be up to 1 to 2 kilobases from the mismatch, is then degraded by an exonuclease to a point beyond the mismatch, without regard to the relative 5' or 3' locations of the nick and mismatch [11]. Resynthesis of the resulting gap replaces the mismatched base(s). |
|
|
| Eukaryotic MMR
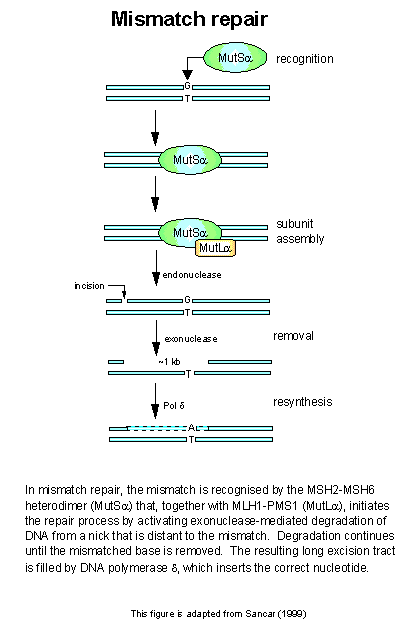
Mismatch repair in eukaryotes
bears a striking resemblance to E. coli methyl-directed MMR, particularly
with respect to mispair specificity and bi-directional excision, however
there are important differences. Although eukaryotes encode several
homologues of both MutS and MutL, no MutH homologue has been identified
(reviewed in [12]), suggesting that another mechanism for strand discimination
may exist. It is currently believed that MMR takes advantage of existing
nicks in the DNA in order to discriminate between the strands (reviewed
in [1, 2]). Following immediately behind the replication fork, MMR
of the lagging strand can take advantage of the gaps between Okazaki fragments
in order to initiate strand degradation and resynthesis. Repair of
the leading strand is believed to be mediated by dissociation of the DNA
polymerase (via a common interaction of the MMR complex and the DNA pold
with PCNA) and degradation of the nascent strand back to the mismatch site
(reviewed in [1, 2]). A second difference between the E. coli model
and that of eukaryotes, is that the yeast and human homologues of MutS
and MutL function as heterodimers comprised of two different MutL (MLH1,
and PMS1 or PMS2) or MutS (MSH2, and MSH3 or MSH6) homologues, rather than
as homodimers. The different pairing combinations appear to the effect
specific activities of the complexes. The MSH2/MSH3 (termed MSHb)
heterodimer has a low affinity for base/base mismatches, but a high affinity
for single stranded loops arising from strand slippage during replication.
This affinity increases with the size of the single stranded loop [13,
14]. In contrast, the MSH2/MSH6 heterodimer (MSHa),
which appears to be the predominant form, exhibits a high affinity for
base/base mismatches, and single stranded loops arising from the insertion/deletion
of only one nucleotide [14].
|
|
|
| MMR
and hereditary colon cancer
Mutations in the MMR genes are believed to account for almost all cases of hereditary nonpolyposis colon cancer (HNPCC). This is one of the most common hereditary cancers, affecting approximately 1 person in 200 and accounting for up to 15% of all colon cancers (reviewed in [12]). Presumably MMR defects result in colon cancer because the cells which line the organ are constantly replicating to replace those sloughed off into the intestinal tract. This high rate of replication is believed to lead to the accumulation of DNA mismatches. Deficiencies in the repair of these mismatches results in the accumulation of mutations and ultimately the development of cancer in these cells. |
|
For more information see: molecular basis of colon cancer HNPCC |
|
|
|
|
| References:
1. Jiricny, J., Eukaryotic mismatch repair: an update. Mutat Res, 1998. 409(3): p. 107-21. 2. Jiricny, J., Replication errors: cha(lle)nging the genome. Embo J, 1998. 17(22): p. 6427-36. 3. Prolla, T.A., DNA mismatch repair and cancer. Curr Opin Cell Biol, 1998. 10(3): p. 311-6. 4. Toft, N.J. and M.J. Arends, DNA mismatch repair and colorectal cancer. J Pathol, 1998. 185(2): p. 123-9. 5. Parker, B.O. and M.G. Marinus, Repair of DNA heteroduplexes containing small heterologous sequences in Escherichia coli. Proc Natl Acad Sci U S A, 1992. 89(5): p. 1730-4. 6. Marinus, M.G. and N.R. Morris, Pleiotropic effects of a DNA adenine methylation mutation (dam-3) in Escherichia coli K12. Mutat Res, 1975. 28(1): p. 15-26. 7. Su, S.S. and P. Modrich, Escherichia coli mutS-encoded protein binds to mismatched DNA base pairs. Proc Natl Acad Sci U S A, 1986. 83(14): p. 5057-61. 8. Grilley, M., et al., Isolation and characterization of the Escherichia coli mutL gene product. J Biol Chem, 1989. 264(2): p. 1000-4. 9. Sancar, A. and J.E. Hearst, Molecular matchmakers. Science, 1993. 259(5100): p. 1415-20. 10. Allen, D.J., et al., MutS mediates heteroduplex loop formation by a translocation mechanism. Embo J, 1997. 16(14): p. 4467-76. 11. Grilley, M., J. Griffith, and P. Modrich, Bidirectional excision in methyl-directed mismatch repair. J Biol Chem, 1993. 268(16): p. 11830-7. 12. Sancar, A., Excision repair invades the territory of mismatch repair. Nat Genet, 1999. 21(3): p. 247-9. 13. Palombo, F., et al., hMutSbeta, a heterodimer of hMsh2 and hMsh3, binds to insertion/deletion loops in DNA. Curr Biol, 1996. 6(9): p. 1181-4. 14. Genschel, J., et al., Isolation of MutSbeta from human cells and
comparison of the mismatch repair specificities of MutSbeta and MutSalpha
[published erratum appears in J Biol Chem 1998 Oct 9;273(41):27034]. J
Biol Chem, 1998. 273(31): p. 19895-901.
|
|
|